Optimality models typically assume:
Haploid or clonal reproduction (sex complicates estimates of fitness, but may not alter predictions)Infinite population size and no subdivision of the population
Strategy set is complete and distinct (i.e., strategies are not subsets of other strategies that are included in the same model)
The optimal solution is genetically possible (e.g., flying elephants!)
Using optimality to understand the economics of prey choice or foraging behavior
First, an example of a discrete optimality model: Choice of two prey with different values (see box 3.2, pg 61 in the text)
Prey type 1 (P1) is worth E1 of energy but takes H1 to break openPrey type 2 (P2) is worth E2 of energy and takes H2 to break open
P1 is more valuable than P2 i.e., E1/H1 > E2/H2Obviously, eat every P1 encountered
The decision to skip or eat P2 will depend on the availability of P1 (which will influence how long it takes to find the next P1 measured as the search time (S1)
A forager should eat P2 when the value of eating P2 is greater that the value of P1, discounted by the time needed to search for the next one: [i.e. E2/H2 > E1/(S1 + H1)]. This leads to the inequality: S1 > (E1H2/E2) - H1
Three predictions emerge from this result:
a) Predator should either: just eat P1 (specialize); or eat both P1 and P2 (generalize)b) Switch should depend only on S1 (availability of P1 ) not the density of P2 (S2)
c) The switch from specialized to generalized feeding should be abrupt
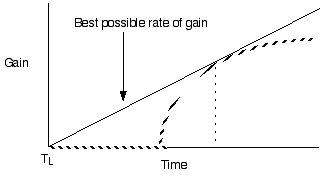
An example of a continuous optimality model: Marginal value analysis of optimal feeding time in food patches (see box 3.1, pg 56 for another application of the marginal value theorum)
Marginal value ideas focus on diminishing returns and the timing of giving up or abandoning a strategy of resource exploitation
1) Assume prey are dispersed in "patches" (some areas better than others)2) Assume the value of patches declines with time (e.g., depletion)
3) Assume patches are initially of equal value
4) Assume patches are distributed evenly (equal travel time between patches)
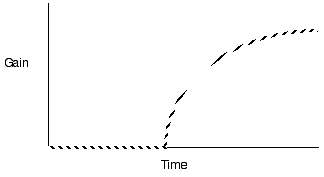
Diminishing return models for single prey types generate decelerating curves through time (measured as cumulative gain or proportion of total gain)
Violate assumption 3: Varying patch quality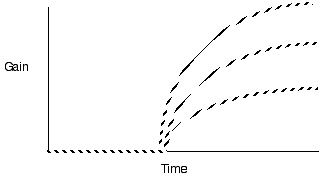
Predict longer residency in the better patches
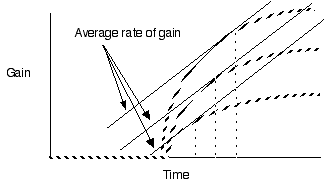
Violate assumption 4: Varying travel time between patches
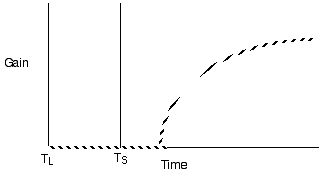
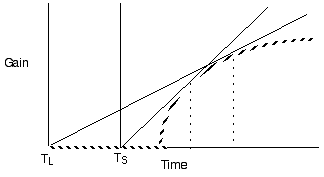
Predict shorter residency when patches are closer together
Other tacit assumptions: perfect knowledge of patch value and distribution... this may explain why animals sometimes stay longer than predicted in bad patches.