| Dept of Biology, Lewis and
Clark College |
|
Dr Kenneth Clifton
|
| |
Biology
352 Lecture Outline
|
|
Introduction
to behavioral ecology and quantitative modeling approaches I:
Optimality
The quest of Behavioral
Ecology: why do animals behave
as they do?
To understand behavioral ecology, we must
understand the link between the expression of behaviors and
individual fitness.
Remember the big three: Food, Sex, and
Death
Diversity in behavior can be examined as a
function of animal Social Organization (where animals are
found and who interacts with whom)
Primary Social Organization:
Behaviors that relate directly to temporal and spatial patterns of
distribution (grouping, spreading out, etc.)
Higher-order Social Organization:
Behaviors that relate to social interactions among animals (mating
systems, parental care, cooperation, etc.)
Why treat dispersion separately from social
behavior issues? After all, most patterns of dispersion involve some
level of social interaction?
Patterns of dispersion (the frequency
and consistency of encounters between individuals) establish the
"template" for social interactions.... selection for "higher
order" social behaviors.
Quantitative approaches to the study of
animal behavior
Why is math important to the study of animal
behavior?
Mathematical methods demand explicit
formulations (in contrast to the vagueness of many verbal
arguments
Models are useful for developing general
concepts that may apply to many different types of
organism
Models may suggest non-intuitive
relationships
Caveats:
Models often rely on overly
simple assumptions,
Testing models can be problematic (are
the data lacking or is the model wrong?)
Hypotheses generated by models may be
neither exclusive, nor exhaustive
Still, mathematical treatments of animal
behavior can be very useful IF you understand the method
behind the madness
Three basic sources of theory in behavioral
ecology:
Comparative methods
Optimality theory
Game theory
Comparative methods can be useful for
identifying generally important aspects of behavior, but theory is
often post hoc.
Overview of quantitative theoretical
approaches to the study of animal behavior
Both optimality and game theory seek to answer
the same question: given a particular ecological scenario (physical
and social environmental conditions) for a given individual, should
it continue a set of behaviors or adopt some alternative?
Definitions:
Strategies: the options available to
an animal (no decision making implied!)
Alternative strategies can either
be discrete or continuous.
Payoffs: the net effect on fitness
associated with a specific strategy (assumes strategies have both
costs and benefits)
Many variables may influence
fitness, though only a subset may be relevant to the
model.
Thus, choice of a "currency" that relates
to fitness is important
Overall
Lifetime Inclusive Fitness (OLIF)
is the only complete currency
It is virtually impossible to
measure
Proxies for OLIF usually relate to one
of the big three (food, sex, or death)
Choosing a modeling approach
Optimality models are useful when we
focus on a single individual in a contest against NATURE (all
environmental variable lumped together)
Such an approach works if none of the
components that make up NATURE respond to an animal's
strategy (i.e. they don't change their behavior or relationship
towards the focal animal and its behavior)
If NATURE responds, theorists often turn to
Game Theory
Optimality theory
Computing payoffs:
Strategies almost universally affect
several components of fitness simultaneously, usually in opposite
ways (because they involve investments of time and energy toward
one thing..... this takes away from another).
Calculating payoffs involves the combination
of various costs and benefits:
We will consider two types of
payoffs: Simple and Conditional
Simple payoffs result from
completely predictable benefits (B) and costs (C).
Animals will often try to
maximize some relationship between B and C (e.g., B - C
or B/C).
Conditional payoffs result when B
& C are fixed, but the probability of experiencing the
benefit or cost is less than 1. There are many
possibilities
1) For example, a strategy
will lead to a benefit (B) with probability
p. If an animal doesn't get B, it gets some
cost (-C) at probability (1 - p).
The expected payoff is:
p(B) +(1-p)(-C)
2) Another example: an animal does
a behavior at cost -C that produces a benefit
(B) with probability p (and no benefit with
probability 1-p). In either case, the animal
accepts the cost (-C)
Now the average expected
payoff is: p(B-C) + (1-p)(-C) = pB - C
Remember, the sum of all
probabilities must equal 1, this helps if there are more
that one type of benefit or cost.
Optimality models typically assume:
Haploid or clonal reproduction (sex
complicates estimates of fitness, but may not alter
predictions)
Infinite population size and no
subdivision of the population
Strategy set is complete and distinct
(i.e., strategies are not subsets of other strategies that are
included in the same model)
The optimal solution is genetically
possible (e.g., if pigs could fly......)
An example of a discrete optimality model:
Choice of two prey with different values
Prey type 1 (T1) is
worth E1 of energy but takes H1 to break
open
Prey type 2 (T2) is worth
E2 of energy and takes H2 to break
open
T1 is more valuable
than T2 i.e., E1/H1 >
E2/H2
Obviously, eat every T1
encountered
The decision to skip or eat
T2 will depend on the availability of
T1 (which will influence how long it takes to
find the next T1 measured as the search time
(S1)
A forager should eat T2
when the value of eating T2 is greater that the
value of T1, discounted by the time needed to
search for the next one: [i.e.
E2/H2 >
E1/(S1 + H1)]. This
leads to the inequality: S1 >
(E1H2/E2) -
H1
Three predictions emerge from this
result:
a) Predator should either:
just eat T1 (specialize); or eat both
T1 and T2 (generalize)
b) Switch should depend only on
S1 (availability of T1 ) not the
density of T2 (S2)
c) The switch from specialized to
generalized feeding should be abrupt
An example of a continuous optimality model:
Marginal value analysis of optimal feeding time in food
patches:
1) Assume prey are dispersed in
"patches" (some areas better than others)
2) Assume the value of patches declines
with time (e.g., depletion)
3) Assume patches are initially of equal
value
4) Assume patches are distributed evenly
(equal travel time between patches)
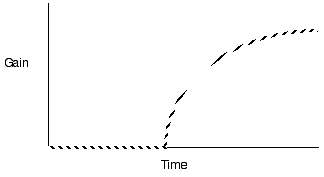
Diminishing return models for single prey
types generate decelerating curves through time
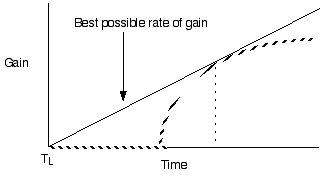
Violate assumption 3: Varying patch
quality
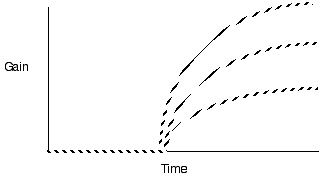
Predict longer residency in the better
patches
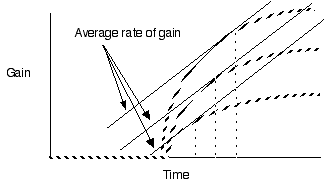
Violate assumption 4: Varying travel time
between patches
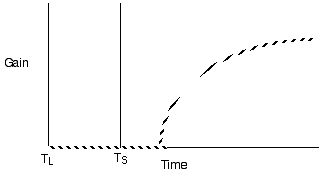
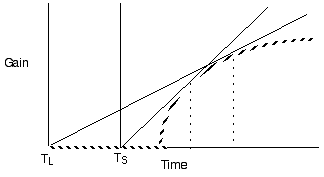
Predict shorter residency when patches are
closer together
Other tacit assumptions: perfect knowledge
of patch value and distribution... this may explain why animals
sometimes stay longer than predicted in bad patches.
Top
Take
me to the previous lecture
Take me to the next lecture
Take me to the course home
page